程序性细胞死亡
Programmed Cell Death (Apoptosis)
凋亡体的形成和caspase的激活
Apoptosome formation and caspase activation
细胞凋亡(Apoptosis)是细胞死亡的一种形式,其利用能量资源以有序或程序化的方式拆除和移除细胞。它在建立正常胚胎发育,维持成体组织稳态和促成多种人类疾病(包括心脏中的某些病理过程)中起重要作用。细胞凋亡由在多细胞生物中保守的独特生化途径介导。细胞凋亡的信号传导从细胞外部(外在或死亡受体途径)或从细胞内部(内在或线粒体途径)开始。在这两种途径中,信号传导导致称为caspase酶家族的激活,其在蛋白水解级联中起作用以拆除和去除垂死细胞。内在死亡途径的激活涉及从线粒体释放细胞色素C和形成凋亡体,凋亡体是激活caspase-9的催化多蛋白平台。
多细胞生物的细胞是高度组织的群落的成员。这个群落的细胞数量受到严格的调控――不仅通过控制细胞分裂的速度,还通过控制细胞的死亡率。如果不再需要细胞,它们就会启动细胞内死亡程序自杀。因此,这一过程被称为程序性细胞死亡(programmed cellular death),尽管它更常见的名称是凋亡(Apoptosis)(源自希腊语,意为“脱落”,如树上的叶子)。
在发育和成年动物组织中发生的凋亡数量可能是惊人的。例如,在正在发育的脊椎动物神经系统中,多达一半或更多的神经细胞通常在形成后不久就会死亡。在一个健康的成年人体内,每小时有数十亿细胞死于骨髓和肠道。如此多的细胞死亡似乎是非常浪费的,尤其是在绝大多数细胞在自杀时是完全健康的。这种大规模细胞死亡的目的是什么?
在某些情况下,答案是明确的。例如,老鼠的爪子是由胚胎发育过程中的细胞死亡塑造的:它们一开始是像铲子一样的结构,单个的手指只有在它们之间的细胞死亡时才会分开(图17-35)。在其他情况下,当细胞形成的结构不再需要时,它们就会死亡。当蝌蚪变成青蛙时,尾巴中的细胞就会死亡,而青蛙不需要的尾巴就会消失(图17-36)。在许多其他情况下,细胞死亡有助于调节细胞数量。例如,在发育中的神经系统中,细胞死亡会调整神经细胞的数量,使之与需要神经刺激的靶细胞数量相匹配。在所有这些病例中,细胞都死于凋亡。
蝌蚪向青蛙蜕变过程中的细胞凋亡。蝌蚪变成青蛙后,蝌蚪尾巴上的细胞被诱导凋亡;结果,尾巴就没了。变形过程中发生的所有变化,包括(更多)
在成人组织中,细胞死亡恰好平衡了细胞分裂。如果不是这样,组织就会生长或收缩。例如,如果成年大鼠的部分肝脏被切除,肝细胞增殖就会增加,以弥补损失。相反,如果给大鼠服用苯巴比妥(苯巴比妥能刺激肝细胞分裂(从而使肝脏增大)),然后停止苯巴比妥治疗,肝细胞凋亡会大大增加,直到肝脏恢复到原来的大小,通常在一周左右的时间内。因此,通过调节细胞死亡率和细胞出生率,肝脏的大小保持不变。
在这一小段中,我们描述了细胞凋亡的分子机制及其控制。在最后一节中,我们将讨论细胞增殖和细胞死亡的细胞外控制如何有助于调节多细胞生物的细胞数量。
细胞凋亡是由细胞内蛋白水解级联介导的
由于急性损伤而死亡的细胞通常会膨胀和破裂。它们会把自己体内的物质溅到周围的邻居身上――这一过程被称为细胞坏死――从而引发潜在的破坏性炎症反应。相比之下,一个经历凋亡的细胞会干净利落地死去,而不会伤害它的邻居。细胞收缩和凝结。细胞骨架崩溃,核膜解体,核DNA分裂成碎片。最重要的是,细胞表面发生了改变,显示了导致濒死细胞在其内容物发生泄漏之前被邻近细胞或巨噬细胞(一种特殊的吞噬细胞,在第24章中讨论过)迅速吞噬的特性(图17-37)。这不仅避免了细胞坏死的破坏性后果,而且还允许死亡细胞的有机成分被摄入细胞回收。
图17-37。细胞死亡。
细胞死亡。这些电子显微照片显示细胞死于(A)坏死或(B和C)凋亡。(A)和(B)中的细胞在培养皿中死亡,而(C)中的细胞在发育中的组织中死亡,并被邻近的细胞吞噬。(更多…)
在所有动物细胞中,负责细胞凋亡的细胞内机制似乎都是相似的。这一机制依赖于一系列蛋白酶,这些蛋白酶在活性位点上有半胱氨酸,并在特定的天冬氨酸上裂解目标蛋白。因此它们被称为caspases。Caspases作为非活性前体或原caspases在细胞中合成,通常被其他Caspases在天冬氨酸裂解激活(图17-38A)。一旦被激活,caspases就会分裂,从而激活其他原caspases,导致蛋白水解级联放大(图17-38B)。一些被激活的caspases会分裂细胞中的其他关键蛋白。例如,一些分裂核薄片,导致核薄片不可逆转的破坏;另一种是分裂一种蛋白质,这种蛋白质通常含有一种不活跃的DNA降解酶(一种DNA酶),释放出DNA酶来切割细胞核中的DNA。这样,细胞迅速而整齐地解体,它的尸体被另一个细胞迅速地吸收和消化。
图17-38。caspase级联参与细胞凋亡。
caspase级联参与细胞凋亡。(A)每一种自杀式蛋白酶都是作为一种不活跃的前酶(原aspase)而产生的,通常由caspase家族的另一成员的蛋白水解裂解激活。如上所示,两个裂开的碎片(更多…)
细胞内细胞死亡途径的激活,就像进入细胞周期的一个新阶段,通常以一种完全的、全有或全无的方式触发。蛋白酶级联不仅具有破坏性和自扩大性,而且是不可逆的,因此一旦细胞在破坏的过程中达到临界点,它就无法逆转。
原caspases是通过与适配器蛋白结合而激活的
所有有核动物细胞都含有自我毁灭的种子,以各种不活跃的原caspases的形式存在,等待着信号来摧毁细胞。因此,caspase的活性在细胞内受到严格的调控,以确保死亡程序在需要时得到控制,这并不奇怪。
如何激活原caspases来启动caspase级联?一个普遍的原理是,这种激活是由适配器蛋白触发的,它将多个特定的原螯合酶(称为引发原螯合酶)以复杂或聚合的形式紧密结合在一起。在某些情况下,引发原螯合酶具有少量的蛋白酶活性,迫使它们结合成一个复合物,导致它们彼此分裂,触发它们的相互激活。在其他情况下,聚合被认为引起构象变化,激活原caspase。瞬间,级联顶部激活的caspase裂解下游的原caspases,放大死亡信号并将其扩散到整个细胞(见图17-38B)。
原 caspase的激活可以通过细胞表面死亡受体的激活从细胞外触发。例如,杀手淋巴细胞(在第24章中讨论)可以通过产生一种叫做Fas配体的蛋白来诱导细胞凋亡,这种配体与靶细胞表面的死亡受体蛋白Fas结合。聚集在一起的Fas蛋白然后招募细胞内的适应蛋白,结合和聚集procaspase-8分子,这些分子分裂并相互激活。活化的caspase-8分子随后激活下游的原螯合酶,诱导细胞凋亡(图17-39A)。一些受到压力或受损的细胞通过同时产生Fas配体和Fas蛋白来杀死自己,从而触发细胞内caspase级联反应。
图17-39。细胞外或细胞内刺激诱导细胞凋亡。
细胞外或细胞内刺激诱导细胞凋亡。(一)细胞外激活。携带Fas配体的杀伤淋巴细胞结合并激活目标细胞表面的Fas蛋白。适配器蛋白与细胞内结合(更多…)
当细胞受到损伤或压力时,它们也可以通过触发原caspase聚集和细胞内的激活来杀死自己。在最容易理解的途径中,线粒体被诱导释放电子载体蛋白细胞色素c(见图14-26)到细胞质中,在那里它结合并激活一种叫做Apaf-1的适配蛋白(图17-39B)。这种原caspase激活的线粒体通路在大多数形式的凋亡中被激活,以启动或加速和放大caspase级联。如前所述,DNA损伤可引发细胞凋亡。这种反应通常需要p53,它可以激活编码蛋白质的基因转录,从而促进细胞色素c从线粒体中释放。这些蛋白质属于Bcl-2家族。
Bcl-2家族蛋白和IAP蛋白是细胞死亡计划的主要细胞内调节因子
细胞内蛋白Bcl-2家族有助于调节原caspases的活化。这个家族的一些成员,如Bcl-2本身或Bcl-XL,通过阻止线粒体释放细胞色素c,至少在一定程度上抑制了细胞凋亡。Bcl-2家族的其他成员不是死亡抑制剂,而是促进原caspase激活和细胞死亡。其中一些细胞凋亡促进因子,如Bad,通过与家族成员的结合和灭活来发挥作用,而另一些,如Bax和Bak,则刺激线粒体释放细胞色素c。如果编码Bax和Bak的基因都是失活的,细胞对大多数诱导凋亡的刺激都具有显著的抵抗能力,这表明这些蛋白在诱导凋亡中起着至关重要的作用。Bax和Bak本身是由Bcl-2家族的其他促凋亡成员(如Bid)激活的。
细胞内凋亡调节因子的另一个重要家族是IAP(凋亡抑制因子)家族。这些蛋白被认为以两种方式抑制细胞凋亡:它们与一些原caspases结合以阻止其激活,它们与caspases结合以抑制其活性。IAP蛋白最初被发现是由某些昆虫病毒产生的蛋白质,这些病毒利用IAP蛋白来阻止受感染的细胞在病毒有时间复制之前杀死自己。当线粒体释放细胞色素c激活Apaf-1时,它们也会释放一种阻断IAPs的蛋白,从而大大提高了死亡激活过程的效率。
细胞内细胞死亡程序也受细胞外信号的调控,细胞外信号可以激活或抑制细胞凋亡。这些信号分子主要通过调节Bcl-2和IAP家族成员的水平或活性发挥作用。我们将在下一节中看到这些信号分子如何帮助多细胞生物调节它们的细胞数量。
总结
在多细胞生物中,不再需要或对生物体构成威胁的细胞会被一种被称为程序性细胞死亡或凋亡的严格调控的细胞自杀过程所破坏。细胞凋亡是由一种叫做caspases的蛋白水解酶介导的,caspases通过分裂细胞质和细胞核中的特定蛋白触发细胞死亡。caspase存在于所有细胞中,作为不活跃的前体,或原caspases,通常被其他Caspases裂解激活,产生蛋白水解caspase级联。激活过程由细胞外或细胞内的死亡信号启动,这导致细胞内的适应分子聚集和激活原caspases。Caspase的激活受Bcl-2和IAP蛋白家族成员的调控。
Apoptosome formation and caspase activation
Apoptosis is a form of cell death which utilizes energy resources to dismantle and remove cells in an orderly or programmed fashion. It plays an essential role in establishing normal embryonic development, maintaining adult tissue homeostasis and contributes to a variety of human diseases including certain pathological processes in the heart. Apoptosis is mediated by a distinct biochemical pathway that is conserved in multicellular organisms. Signaling for apoptosis is initiated from outside the cell (extrinsic or death receptor pathway) or from inside the cell (intrinsic or mitochondrial pathway). In both pathways, signaling results in the activation of a family of cysteine proteases, named caspases, that act in a proteolytic cascade to dismantle and remove the dying cell. The activation of the intrinsic death pathway involves the release of cytochrome c from the mitochondria and formation of the apoptosome, a catalytic multiprotein platform that activates caspase-9. There is evidence that the mitochondrial pathway is involved in ischemia-induced myocyte apoptosis in the heart. Diminished expression of pro-apoptotic factors and/or expression of certain inhibitors of the apoptosome may raise the threshold for apoptosis in long-lived post-mitotic cells including myocytes of the heart.
The cells of a multicellular organism are members of a highly organized community. The number of cells in this community is tightly regulated―not simply by controlling the rate of cell division, but also by controlling the rate of cell death. If cells are no longer needed, they commit suicide by activating an intracellular death program. This process is therefore called programmed cell death, although it is more commonly called apoptosis (from a Greek word meaning “falling off,” as leaves from a tree).
The amount of apoptosis that occurs in developing and adult animal tissues can be astonishing. In the developing vertebrate nervous system, for example, up to half or more of the nerve cells normally die soon after they are formed. In a healthy adult human, billions of cells die in the bone marrow and intestine every hour. It seems remarkably wasteful for so many cells to die, especially as the vast majority are perfectly healthy at the time they kill themselves. What purposes does this massive cell death serve?
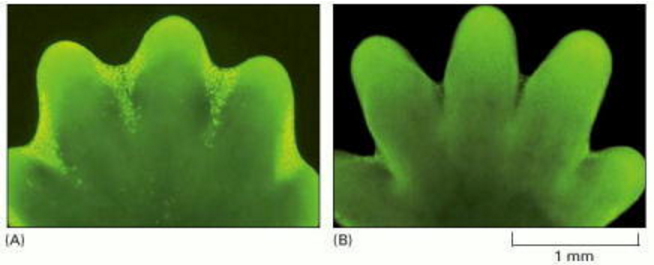
In some cases, the answers are clear. Mouse paws, for example, are sculpted by cell death during embryonic development: they start out as spadelike structures, and the individual digits separate only as the cells between them die (Figure 17-35). In other cases, cells die when the structure they form is no longer needed. When a tadpole changes into a frog, the cells in the tail die, and the tail, which is not needed in the frog, disappears (Figure 17-36). In many other cases, cell death helps regulate cell numbers. In the developing nervous system, for example, cell death adjusts the number of nerve cells to match the number of target cells that require innervation. In all these cases, the cells die by apoptosis.
Figure 17-35. Sculpting the digits in the developing mouse paw by apoptosis.
Figure 17-35
Sculpting the digits in the developing mouse paw by apoptosis. (A) The paw in this mouse embryo has been stained with a dye that specifically labels cells that have undergone apoptosis. The apoptotic cells appear as bright green dots between the developing (more...)
Figure 17-36. Apoptosis during the metamorphosis of a tadpole into a frog.
Figure 17-36
Apoptosis during the metamorphosis of a tadpole into a frog. As a tadpole changes into a frog, the cells in the tadpole tail are induced to undergo apoptosis; as a consequence, the tail is lost. All the changes that occur during metamorphosis, including (more...)
In adult tissues, cell death exactly balances cell division. If this were not so, the tissue would grow or shrink. If part of the liver is removed in an adult rat, for example, liver cell proliferation increases to make up the loss. Conversely, if a rat is treated with the drug phenobarbital―which stimulates liver cell division (and thereby liver enlargement)―and then the phenobarbital treatment is stopped, apoptosis in the liver greatly increases until the liver has returned to its original size, usually within a week or so. Thus, the liver is kept at a constant size through the regulation of both the cell death rate and the cell birth rate.
In this short section, we describe the molecular mechanisms of apoptosis and its control. In the final section, we consider how the extracellular control of cell proliferation and cell death contributes to the regulation of cell numbers in multicellular organisms.
Apoptosis Is Mediated by an Intracellular Proteolytic Cascade
Cells that die as a result of acute injury typically swell and burst. They spill their contents all over their neighbors―a process called cell necrosis―causing a potentially damaging inflammatory response. By contrast, a cell that undergoes apoptosis dies neatly, without damaging its neighbors. The cell shrinks and condenses. The cytoskeleton collapses, the nuclear envelope disassembles, and the nuclear DNA breaks up into fragments. Most importantly, the cell surface is altered, displaying properties that cause the dying cell to be rapidly phagocytosed, either by a neighboring cell or by a macrophage (a specialized phagocytic cell, discussed in Chapter 24), before any leakage of its contents occurs (Figure 17-37). This not only avoids the damaging consequences of cell necrosis but also allows the organic components of the dead cell to be recycled by the cell that ingests it.
Figure 17-37. Cell death.
Cell death. These electron micrographs show cells that have died by (A) necrosis or (B and C) apoptosis. The cells in (A) and (B) died in a culture dish, whereas the cell in (C) died in a developing tissue and has been engulfed by a neighboring cell. (more...)
The intracellular machinery responsible for apoptosis seems to be similar in all animal cells. This machinery depends on a family of proteases that have a cysteine at their active site and cleave their target proteins at specific aspartic acids. They are therefore called caspases. Caspases are synthesized in the cell as inactive precursors, or procaspases, which are usually activated by cleavage at aspartic acids by other caspases (Figure 17-38A). Once activated, caspases cleave, and thereby activate, other procaspases, resulting in an amplifying proteolytic cascade (Figure 17-38B). Some of the activated caspases then cleave other key proteins in the cell. Some cleave the nuclear lamins, for example, causing the irreversible breakdown of the nuclear lamina; another cleaves a protein that normally holds a DNA-degrading enzyme (a DNAse) in an inactive form, freeing the DNAse to cut up the DNA in the cell nucleus. In this way, the cell dismantles itself quickly and neatly, and its corpse is rapidly taken up and digested by another cell.
Figure 17-38. The caspase cascade involved in apoptosis.
The caspase cascade involved in apoptosis. (A) Each suicide protease is made as an inactive proenzyme (procaspase), which is usually activated by proteolytic cleavage by another member of the caspase family. As indicated, two of the cleaved fragments (more...)
Activation of the intracellular cell death pathway, like entry into a new stage of the cell cycle, is usually triggered in a complete, all-or-none fashion. The protease cascade is not only destructive and self-amplifying but also irreversible, so that once a cell reaches a critical point along the path to destruction, it cannot turn back.
Procaspases Are Activated by Binding to Adaptor Proteins
All nucleated animal cells contain the seeds of their own destruction, in the form of various inactive procaspases that lie waiting for a signal to destroy the cell. It is therefore not surprising that caspase activity is tightly regulated inside the cell to ensure that the death program is held in check until needed.
How are procaspases activated to initiate the caspase cascade? A general principle is that the activation is triggered by adaptor proteins that bring multiple copies of specific procaspases, known as initiator procaspases, close together in a complex or aggregate. In some cases, the initiator procaspases have a small amount of protease activity, and forcing them together into a complex causes them to cleave each other, triggering their mutual activation. In other cases, the aggregation is thought to cause a conformational change that activates the procaspase. Within moments, the activated caspase at the top of the cascade cleaves downstream procaspases to amplify the death signal and spread it throughout the cell (see Figure 17-38B).
Procaspase activation can be triggered from outside the cell by the activation of death receptors on the cell surface. Killer lymphocytes (discussed in Chapter 24), for example, can induce apoptosis by producing a protein called Fas ligand, which binds to the death receptor protein Fas on the surface of the target cell. The clustered Fas proteins then recruit intracellular adaptor proteins that bind and aggregate procaspase-8 molecules, which cleave and activate one another. The activated caspase-8 molecules then activate downstream procaspases to induce apoptosis (Figure 17-39A). Some stressed or damaged cells kill themselves by producing both the Fas ligand and the Fas protein, thereby triggering an intracellular caspase cascade.
Figure 17-39. Induction of apoptosis by either extracellular or intracellular stimuli.
Induction of apoptosis by either extracellular or intracellular stimuli. (A) Extracellular activation. A killer lymphocyte carrying the Fas ligand binds and activates Fas proteins on the surface of the target cell. Adaptor proteins bind to the intracellular (more...)
When cells are damaged or stressed, they can also kill themselves by triggering procaspase aggregation and activation from within the cell. In the best understood pathway, mitochondria are induced to release the electron carrier protein cytochrome c (see Figure 14-26) into the cytosol, where it binds and activates an adaptor protein called Apaf-1 (Figure 17-39B). This mitochondrial pathway of procaspase activation is recruited in most forms of apoptosis to initiate or to accelerate and amplify the caspase cascade. DNA damage, for example, as discussed earlier, can trigger apoptosis. This response usually requires p53, which can activate the transcription of genes that encode proteins that promote the release of cytochrome c from mitochondria. These proteins belong to the Bcl-2 family.
Bcl-2 Family Proteins and IAP Proteins Are the Main Intracellular Regulators of the Cell Death Program
The Bcl-2 family of intracellular proteins helps regulate the activation of procaspases. Some members of this family, like Bcl-2 itself or Bcl-XL, inhibit apoptosis, at least partly by blocking the release of cytochrome c from mitochondria. Other members of the Bcl-2 family are not death inhibitors, but instead promote procaspase activation and cell death. Some of these apoptosis promoters, such as Bad, function by binding to and inactivating the death-inhibiting members of the family, whereas others, like Bax and Bak, stimulate the release of cytochrome c from mitochondria. If the genes encoding Bax and Bak are both inactivated, cells are remarkably resistant to most apoptosis-inducing stimuli, indicating the crucial importance of these proteins in apoptosis induction. Bax and Bak are themselves activated by other apoptosis-promoting members of the Bcl-2 family such as Bid.
Another important family of intracellular apoptosis regulators is the IAP (inhibitor of apoptosis) family. These proteins are thought to inhibit apoptosis in two ways: they bind to some procaspases to prevent their activation, and they bind to caspases to inhibit their activity. IAP proteins were originally discovered as proteins produced by certain insect viruses, which use them to prevent the infected cell from killing itself before the virus has had time to replicate. When mitochondria release cytochrome c to activate Apaf-1, they also release a protein that blocks IAPs, thereby greatly increasing the efficiency of the death activation process.
The intracellular cell death program is also regulated by extracellular signals, which can either activate apoptosis or inhibit it. These signal molecules mainly act by regulating the levels or activity of members of the Bcl-2 and IAP families. We see in the next section how these signal molecules help multicellular organisms regulate their cell numbers.
Summary
In multicellular organisms, cells that are no longer needed or are a threat to the organism are destroyed by a tightly regulated cell suicide process known as programmed cell death, or apoptosis. Apoptosis is mediated by proteolytic enzymes called caspases, which trigger cell death by cleaving specific proteins in the cytoplasm and nucleus. Caspases exist in all cells as inactive precursors, or procaspases, which are usually activated by cleavage by other caspases, producing a proteolytic caspase cascade. The activation process is initiated by either extracellular or intracellular death signals, which cause intracellular adaptor molecules to aggregate and activate procaspases. Caspase activation is regulated by members of the Bcl-2 and IAP protein families.
By agreement with the publisher, this book is accessible by the search feature, but cannot be browsed.
source:
Molecular Biology of the Cell. 4th edition.
Alberts B, Johnson A, Lewis J, et al.
New York: Garland Science; 2002.
Copyright © 2002, Bruce Alberts, Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walter; Copyright © 1983, 1989, 1994, Bruce Alberts, Dennis Bray, Julian Lewis, Martin Raff, Keith Roberts, and James D. Watson .
Programmed Cell Death (Apoptosis) - Molecular Biology of the Cell - NCBI Bookshelf https://www.ncbi.nlm.nih.gov/books/NBK26873/
Absolute Calibration of Mitochondrial Membrane Potential https://www.imageanalyst.net/calibrationwizard.html
https://www.sciencedirect.com/science/article/pii/S002228280400118X
.png)
.png)
.png)
.png)
.png)
.png)
.png)
.png)